Шишковидная железа третий глаз, Эпифиз сквозь время
У волнистого попугайчика она имеет вид пузырька, соединённого с эпифизом, а у лебедя даже отделяется от него []. Задняя доля гипофиза относится к промежуточному мозгу , однако передняя его доля представляет собой железу внутренней секреции. В XIX и начале XX века были популярны предположения и утверждения [54] о гомологии теменного глаза позвоночных медианному глазу оболочников и даже некоторых членистоногих , основанные на расположении этих глаз и неинвертированности их сетчатки [] [] [] [] []. Более менее познавательно, спс. Пинеальный комплекс — основная после парных глаз светочувствительная структура низших позвоночных.
В самом узком смысле это название относится только к хорошо развитому парапинеальному органу рептилий [18] [19] , а в самом широком — и к светочувствительным органам пинеального комплекса некоторых земноводных [20] [21] , рыб [22] и бесчелюстных [23] [24]. У остальных современных позвоночных оба этих органа не глазоподобны, а часто и сильно редуцированы , и их не называют глазами.
Так, у млекопитающих и птиц пинеальный орган скрыт под черепом и часто в глубине мозга , а парапинеальный исчез совсем. Некоторые животные потеряли и тот и другой [15] [27]. К глазоподобным производным промежуточного мозга применяется множество терминов, которые у разных авторов часто имеют разный смысл [28] [21]. Пинеальный и парапинеальный орган составляют пинеальный комплекс [29] [30] , также известный как эпифизарный комплекс [20]. Иногда оба органа этого комплекса называют пинеальными [31].
Пинеальный орган в узком смысле лат. Парапинеальный орган лат. Кроме того, он известен как теменной париетальный орган [35] [32] [30] лат.
Если пинеальный или парапинеальный орган имеет более или менее глазоподобный вид, его называют соответственно « пинеальный глаз » [42] [43] или « парапинеальный глаз » [27] , но иногда пинеальным глазом называют любой из них [36]. Кроме того, и тот и другой известен как третий глаз или непарный глаз [12] [24] [44] , а также «медианный глаз» [45] и «эпифизарный глаз» [15] [46] [36].
Если эти органы не глазоподобны, их называют железами [32] лат. Термин « теменной глаз » тождественно « париетальный глаз » [33] [47] , англ. По другим представлениям третий глаз бесхвостых — не парапинеальный орган, а часть пинеального [16].
Расположен он в передней части головы. В связи с этим его часто называют « лобный глаз » « фронтальный глаз », « фронтальный орган » [49] [33] [50]. Теменной парапинеальный глаз рептилий изредка называют « орган Лейдига » или «лейдигов орган» [51] [46] [52] , хотя такое же название носит и ряд других органов разных животных.
Названия «теменной глаз» [53] [54] , «лобный глаз» [55] , «фронтальный глаз» [56] , «медианный глаз» [55] , «непарный глаз» [56] , « третий глаз » тоже относятся не только к данным органам: их применяют, в частности, к простым глазкам членистоногих. Эмбриогенез пинеального комплекса, как и сетчатки парных глаз, прослежен начиная от стадии нервной пластинки , и установлено, что они развиваются из её соседних областей [3].
Зачатки пинеального и парапинеального органа представляют собой выпячивания крыши третьего желудочка головного мозга , которая входит в состав эпиталамуса — структуры в промежуточном мозгу. Таким образом, полость этих зачатков вначале сообщается с третьим желудочком и заполнена спинномозговой жидкостью [57].
У взрослых животных многих видов особенно у высших позвоночных это сообщение, а иногда и сама полость, теряется [58] [20] [59]. Пинеальный комплекс — одна из самых изменчивых структур мозга позвоночных [60] : в разных группах его строение отличается очень сильно. Если парапинеальный орган возникает, то его зачаток расположен спереди и слева от зачатка пинеального [61] но есть данные, что у некоторых рептилий они развиваются из одного общего выпячивания [26] [62] [16].
У взрослых рептилий парапинеальный орган находится выше пинеального, а у миног и костистых рыб — ниже [59]. Асимметрия пинеального комплекса проявляется не только в его морфологии, но и в организации нейронов , нейрохимии и в связях с другими частями мозга.
От парапинеального переднего и левого органа нервные волокна идут к левой стороне поводка [en] крупной структуры в эпиталамусе [16] , а от пинеального заднего и правого — к правой или ни к какой.
Это указывает на то, что у предков позвоночных органы пинеального комплекса образовывали симметричную пару [63] [32] [15] [27] [31]. Поводок у большинства позвоночных тоже асимметричен, причём его асимметрия чаще всего соответствует асимметрии пинеального комплекса если таковая есть [63] [16] [64].
У многих бесчелюстных у круглоротых и костнопанцирных [65] [66] и большинства костных рыб лучше развита правая часть поводка, а у хрящевых рыб , латимерии , амфибий и рептилий — левая [16] [67].
У некоторых животных его асимметричное развитие — результат влияния парапинеального органа [64] [68] [69]. Два выроста мозга, расположенные рядом с пинеальным комплексом, — парафиз лат. Пинеальный комплекс — основная после парных глаз светочувствительная структура низших позвоночных. Кроме него и парных глаз, специализированные фоторецепторные клетки есть только в некоторых областях мозга около желудочков хотя чувствительность к свету могут проявлять и некоторые клетки других органов, в том числе кожи [31] [29].
У некоторых животных часть пинеального комплекса принимает вид хорошо обособленного округлого пузырька с явными адаптациями к светочувствительной функции [2] — непарного глаза. Ножка, которая соединяет его с мозгом на ранних стадиях развития, может сохраняться у миног [73] [74] и немногих ящериц [75] или исчезать у бесхвостых земноводных , большинства ящериц и гаттерии [76]. Впоследствии из этого глаза прорастает нерв , который соединяет его с мозгом заново [39] [77] [18] с возрастом часто исчезает и он [78] [79] [80].
У рептилий глазоподобный вид принимает парапинеальный орган, а у миног — и пинеальный, и парапинеальный. У бесхвостых земноводных непарный глаз развивается из пузырька, расположенного, вероятно [16] , с задней стороны зачатка пинеального органа [19] по другим данным — с передней [78].
Это а также характер его нервных соединений указывает на то, что он не гомологичен парапинеальному органу [16] [19]. Обычно его считают частью пинеального органа [25] [16] [19] [18] [81] [32] , иногда — парапинеальным органом [21] [20] [26]. Между начальными стадиями онтогенеза парных и непарных глаз есть много общего.
Эксперименты с жабами показали, что зачаток пинеального органа в некоторых условиях способен развиться в глаз, почти идентичный парным глазам. Это наблюдалось при его пересадке головастикам от головастиков более раннего возраста, а также при удалении у них парных глаз [44].
Но в норме строение парных и непарных глаз сильно различается. В ходе развития непарного глаза, в отличие от парных, не происходит впячивания его передней стенки внутрь и превращения её в сетчатку.
Вместо этого она остаётся прозрачной, а у некоторых рептилий [79] даже становится двояковыпуклой. Часто её называют хрусталиком [12] [27] [82]. Таким образом, хрусталик непарного глаза образуется из нервной эктодермы [15] , а не из кожной, и поэтому не гомологичен хрусталику парных [83]. Клетки его внутренней стороны несут реснички — свидетельство его происхождения от стенки нервной трубки [84] [85] [15]. Этот хрусталик может быть не только двояковыпуклым или плоско-выпуклым, но и выпукло-вогнутым, и даже двояковогнутым [79].
К аккомодации непарный глаз не способен [86]. Сетчатка непарного глаза образуется из задней стенки его зачатка, а не из передней. Как следствие, она всегда [15] [23] неинвертированная.
В парных глазах позвоночных сетчатка всегда инвертирована светочувствительные клетки находятся с её задней стороны, их фоторецепторные сегменты направлены также назад, и, таким образом, свет по пути к ним проходит через несколько дополнительных слоёв клеток. Задняя стенка зачатка парного глаза даёт только самый задний слой его сетчатки — пигментный эпителий.
Ему и гомологична сетчатка непарного глаза [87]. Среди современных животных лучше всего развитый третий глаз имеют некоторые ящерицы и гаттерия. Похожее строение имеют и оба непарных глаза миног. У этих животных лучше развит пинеальный орган. Парапинеальный, лежащий ниже вплотную к мозгу и немного левее, отличается от него меньшим размером и более простым строением: его верхняя стенка слабее отличается от нижней [63] [13] [33] [90] , а светочувствительные и ганглионарные клетки в нём более редки [23] [16] [41].
Если теменной глаз слабо развит, его стенки как и стенки неглазоподобного пинеального органа не дифференцированы на хрусталик и сетчатку. Такой вид он имеет, например, у ряда хамелеонов , агамовых и сцинковых. Кроме того, у них сохраняется ножка, соединяющая его с пинеальным органом [91] [92] [93] [75].
Непарный глаз миног, земноводных и рептилий обычно круглый или приплюснутый, а у некоторых ящериц — вертикально вытянутый [79]. Пинеальный орган современных рыб имеет трубкоподобную или мешковидную форму и менее глазоподобен [94] [95].
Тем не менее у тех рыб, у которых его приспособления к светочувствительности ярко выражены, его тоже называют теменным, медианным или третьим глазом [44] [22] [15] [24] [96]. У карповых рыб из рода Labeo его часть образует обособленный пузырёк, лежащий в специальном отверстии в черепе под прозрачным участком кожи и, таким образом, напоминает теменной глаз ящериц [38].
Размер непарного глаза современных бесчелюстных, амфибий и рептилий составляет несколько десятых долей миллиметра. У 1,8-метрового гигантского варана его длина вдоль оптической оси равна 0,4 мм; такой же размер он имеет и у сантиметровой гаттерии [97] [80]. Диаметр теменного глаза зелёной игуаны составляет 0,5—0,8 мм [98]. У других современных пресмыкающихся он ещё меньше [80]. У бесхвостых земноводных его размер обычно лежит в пределах 0,10—0,15 мм [99].
У миног диаметр пинеального глаза равен 0,4—0,5 мм, а парапинеального — в 1,5—2 раза меньше [13] [] [23]. Теменной глаз многих вымерших животных, судя по размеру их теменного отверстия, был во много раз больше [] []. Это даёт основания предполагать его более сложное устройство, но явных признаков этого на окаменелостях нет [80]. С другой стороны, есть версия, что неровности на стенках теменного отверстия ихтиозавров и плезиозавров могут быть следами глазодвигательных мышц [] , которых у непарного глаза современных животных нет [] [79] [11].
Обычно теменной глаз лучше развит у молодых животных; с возрастом он в большей или меньшей степени атрофируется. Часто у взрослых особей исчезает и его нерв [26] [79] [80] , а изредка и соответствующее отверстие в черепе это известно у некоторых ящериц [80] [] [92] , ископаемых синапсид [] и остракодерм []. С возрастом обычно деградируют непарные глаза и миног [13] , и земноводных [] , и рептилий [15] [93] , а также менее глазоподобный парапинеальный орган рыб [] [14].
Сетчатка непарного глаза состоит из клеток тех же трёх типов, которые есть и в неглазоподобных органах пинеального комплекса: пинеалоцитов [en] , ганглионарных нейронов и поддерживающих клеток [] [] [87] [] [94] [95]. Фоторецепция, синтез мелатонина и поддержание эндогенного суточного ритма его интенсивности происходит в пинеалоцитах [3].
Они очень близки по строению, функциям и происхождению к светочувствительным клеткам парных глаз — палочкам и колбочкам [] [87] [3]. Признаки этих типов клеток в них смешаны [] [31] фоторецепторы парных глаз чётко делятся на палочки и колбочки у всех челюстноротых []. Они могут делиться на разные типы по морфологии и спектральной чувствительности [41]. Так, есть признаки наличия в непарном глазе амфибий, как и в парных, двойных фоторецепторов наряду с одинарными [].
В органах пинеального комплекса, которые находятся под черепом, преобладают палочкоподобные пинеалоциты с механизмом восприятия света, характерным для ночного зрения , а во внечерепных непарных глазах — колбочкоподобные [31]. У низших позвоночных пинеалоциты намного больше похожи на фоторецепторы парных глаз, чем у высших. Как и колбочки и палочки парных глаз, пинеалоциты имеют внутренний и наружный сегмент, соединённые своеобразной неподвижной ресничкой.

В отличие от обычных ресничек в том числе ресничек прочих клеток сетчатки и хрусталика теменного глаза [85] , в ней нет центральных микротрубочек []. Иногда этих ресничек в клетке две, но чаще одна [31]. Она продолжается в наружный сегмент пинеалоцита, сформированный стопкой крепящихся к ней дисков, образованных складками клеточной мембраны.
В этих дисках и осуществляется фоторецепция. Они постоянно обновляются, а сброшенные фагоцитируются специальными клетками иногда — внутренними сегментами самих же пинеалоцитов [] []. Тем не менее у земноводных и миног но не у ящериц полость непарного глаза засорена множеством сброшенных дисков []. Длина наружного сегмента пинеалоцита достигает 10—20 мкм, а количество дисков в нём — нескольких сотен, а иногда и тысяч []. У млекопитающих и птиц этот сегмент обычно сильно редуцирован [31]. Как и фоторецепторы парных глаз, пинеалоциты воспринимают свет с помощью различных опсинов , в частности, родопсина [6] [31] [] , и нуждаются в витамине A [89].
В непарных глазах найдены и такие опсины, которых нет в парных; некоторые из них относятся к отдельным семействам этих белков [6] [] [] [] []. Пинеалоциты соединены синапсами с ганглионарными нейронами , аксоны которых и образуют нерв, идущий к мозгу []. Этим сетчатка непарных глаз отличается от сетчатки парных, где у большинства позвоночных фоторецепторы соединяются с ганглионарными нейронами не напрямую, а через ряд промежуточных клеток биполярные , горизонтальные и амакриновые нейроны [34] [] [].
С каждой ганглионарной клеткой соединяется несколько пинеалоцитов, и поэтому волокон в нерве теменного глаза намного меньше, чем фоторецепторов в его сетчатке [].
У миног [41] , рыб и млекопитающих [] [] в составе нерва пинеального органа есть и аксоны самих пинеалоцитов. Кроме пинеалоцитов и ганглионарных нейронов, в сетчатке теменного глаза есть поддерживающие клетки, разбросанные между фоторецепторами. Эти клетки обычно содержат тёмный пигмент предположительно меланин [].
У ящериц этого пигмента много [79] [] , а его гранулы могут двигаться вверх и вниз и таким образом обеспечивать адаптацию глаза к изменениям освещённости [] как и в парных глазах многих позвоночных []. У лягушек его меньше [88] , а у миног чаще всего нет вообще [] []. Но у них в этих клетках есть блестящие микрокристаллики по всей видимости, гуанина , которые придают непарным глазам миног серебристо-белый цвет [23] [63] [13] [].
Похожие кристаллики придают серебристый блеск и теменному глазу ящериц, но у них они находятся в клетках, окружающих его снаружи. Количество этого светлого пигмента у ящериц [] [87] , как и тёмного пигмента у миног [] , может сильно варьировать в пределах вида. Нерв непарного глаза земноводных и рептилий обычно труднозаметен а часто и исчезает с возрастом [78] [26] [79] [80] и во многих старых работах [33] написано, что он отсутствует [] [27] [].
Например, у пустынной игуаны этот нерв имеет толщину всего 15—25 мкм при количестве волокон от до []. У других изученных ящериц он обычно тоже составлен несколькими сотнями волокон, а у лягушек — несколькими десятками [] [8].
Максимальное зарегистрированное число аксонов в этом нерве — около []. У ящериц и миног он иногда поделён на несколько нитей [79] [97] []. Большинство его волокон безмиелиновые [] [] [88] миелинизированы только отростки клеток с ахроматическим откликом см. У миног и земноводных его по ряду соображений иногда называют не нервом, а трактом [] [88] []. Но по строению он больше похож на периферические нервы, чем на тракты в ЦНС [8] [].
У миног — единственных современных животных, у которых оба органа пинеального комплекса относительно глазоподобны — они иннервируют разные области мозга. От парапинеального органа волокна идут через коммиссуру поводков [en] в левую часть поводка [en] , паллиум конечного мозга , предкрышечное поле [en] и ядра в среднем мозгу и передней части промежуточного мозга [19]. Пинеальный орган иннервирует другие зоны в среднем и промежуточном мозгу — его нерв идёт через заднюю коммиссуру [en] в правую часть поводка [31] [15] [32] [63] [73] , оптический тектум , покрышку среднего мозга , ретикулярную формацию и ядро глазодвигательного нерва [19].
Но в других исследованиях иннервация поводка непарными глазами у миног не наблюдалась [41]. У хрящевых и костных рыб нервные волокна от пинеального органа идут во многие области среднего и промежуточного мозга [19] , в частности, в заднюю коммиссуру, поводковые ядра, предкрышечное поле, околожелудочковое серое вещество , покрышку среднего мозга и, у некоторых видов, в преоптическую зону [en] [] [29] и оптический тектум [].
У амфибий и рептилий нерв непарного глаза идёт к внутричерепной части пинеального комплекса и вливается в пинеальный тракт, соединяющий её с промежуточным мозгом. Составляя в этом тракте всего несколько процентов волокон [8] [29] [] , аксоны клеток этого глаза направляются дальше. Области мозга, где они кончаются, у земноводных и рептилий существенно отличаются [64] [16] , но и у тех и у других примерно совпадают с областями, иннервируемыми внутричерепным пинеальным органом [] [29].
У тех животных, у которых непарного глаза нет — млекопитающих и птиц — почти нет и волокон, идущих в мозг из пинеального органа [29] [16] [19]. У земноводных и рептилий аксоны из непарного глаза проходят через заднюю коммиссуру [58] [26] по другим данным — мимо неё [64] , после чего расходятся по сторонам и вниз [] и направляются в различные структуры промежуточного , среднего , а иногда и конечного мозга.
Эта иннервация, как и сам поводок, примечательна своей сильной асимметрией у многих позвоночных из разных классов. Теменной глаз рептилий, как правило, соединён с левой половиной этой структуры [16] [31] [64] [] [29].
Данные о его связи у некоторых ящериц с правой половиной [15] [62] [33] , возможно, ошибочны [] [64] , а возможно, объясняются тем, что этот нерв вырастает из глаза уже после его отпочковывания от мозга и не имеет отношения к их первичному соединению [].
У бесхвостых земноводных волокна непарного глаза по одним данным тоже идут, среди прочего, в левую половину поводка [19] которая у них, как и у рептилий, больше правой [16] [] , а по другим — симметрично расходятся в другие области мозга [16] [64].
Остальные идут в миндалевидные тела , предкрышечное поле и околожелудочковое серое вещество [64] [29] []. Аксоны клеток теменного глаза ящериц направляются, кроме поводка, в предкрышечное поле, дорсолатеральное ядро таламуса и преоптические зоны гипоталамуса и конечного мозга [19] [29] [82].
Определение мест, где кончаются нервные волокна органов пинеального комплекса, — трудная задача, и здесь остаётся много неясного [] [14] [29]. Есть данные, что теменной глаз ящериц не иннервирует зрительные области мозга [6] [].
Тем не менее места окончания его волокон как и у миног [41] частично перекрываются с местами окончания волокон зрительных нервов парных глаз [29] [19] []. В частности, многие аксоны от парных и непарных глаз кончаются в предкрышечном поле, и предполагается, что оно играет важную роль в обработке поступающей от них информации [4].
У животных, у которых органы пинеального комплекса развиваются до глазоподобного вида, волокон, идущих туда из мозга, нет или почти нет [] []. Но у некоторых, в том числе миног, рыб [57] [] , земноводных и ящериц [] [] [19] [64] , они известны. Эфферентная иннервация этих органов хорошо развита только у млекопитающих и птиц, у которых они не глазоподобны.
У них пинеальный орган иннервируют симпатические нервные волокна из верхнего шейного ганглия [en] [19] , к которому идут через ряд разных структур нервные пути из супрахиазматического ядра гипоталамуса. Оно служит водителем суточных ритмов организма биологическими часами и управляет ритмом синтеза мелатонина в пинеальном органе. У птиц роль этого ядра в регуляции суточных ритмов меньше, чем у млекопитающих, и иннервация пинеального органа развита слабее [] [3] []. У миног и амниот эфферентная иннервация пинеального комплекса существенно отличается, и это указывает на то, что она у них возникла независимо [19].
Третий глаз всегда лежит под кожей, а иногда и под черепом, но у многих животных кожа над ним относительно прозрачна. Такой просвечивающий участок кожи встречается даже у тех животных, у которых структура органов пинеального комплекса далека от глазоподобной — например, у некоторых черепах и многих рыб.
Он известен как пинеальное окно [] или теменное пятно [30] англ. У земноводных его называют также пятном Штиды или лобным пятном [50] англ. У рыб пигментный слой в области пинеального органа может присутствовать, но лежать ниже него — между ним и мозгом например, у сардин [] []. У некоторых окунеобразных — скумбриевых в частности, тунца , марлиновых и меч-рыбы — ткани, проводящие свет к этому органу, образуют сложную структуру, известную как пинеальный аппарат.
Он начинается хорошо заметным на лбу пинеальным окном, под которым находится отверстие между лобными костями, затянутое хрящевой тканью.
Вокруг него отростки костей образуют идущую вглубь трубку, в которой лежит пинеальный орган. Весь пинеальный аппарат погружён в жировую ткань [89] []. Непарный глаз ящериц лежит под большой чешуйкой, центральная часть которой более или менее прозрачна и часто слегка выпукла [93].
У некоторых ящериц например, у варанов пинеальное окно — самая заметная деталь на верхней стороне головы. У гаттерии в молодом возрасте оно не покрыто чешуёй и тоже хорошо заметно.
Позже на него надвигаются соседние чешуйки, и у взрослых гаттерий на положение теменного глаза указывает только ослабленная пигментация кожи [] []. Но у кожистой черепахи , не имеющей ни теменного глаза, ни теменного отверстия, хорошо заметное пинеальное окно сохраняется всю жизнь [] []. У гаттерии между кожей и теменным глазом лежит прозрачная линзоподобная структура из соединительной ткани , которая занимает верхнюю часть теменного отверстия []. Похожая структура есть и у некоторых сомообразных рыб [] , а также у миног у которых она лежит между кожей и черепом [63] [13].
Такие прозрачные ткани над теменным глазом иногда называют его роговицей [12] [6] [40]. Аналогов радужки и век у непарных глаз нет [10] [11] , но у некоторых рыб например, у нерки [] доступ света к пинеальному органу регулируется сокращением и растягиванием пигментных клеток [15] [] []. У акул прозрачность кожи в этом месте может быть всемеро больше обычной [29]. Если в рот лимонной акулы поместить лампочку, через пинеальное окно её свет хорошо виден снаружи []. Но достаточное для влияния на физиологические процессы [8] количество света может проходить и сквозь череп животных, у которых нет ни пинеального окна, ни отверстия в черепе [29] [57] [] в том числе некоторых млекопитающих [] [31] [] [8] и птиц [] [29].
У многих позвоночных в черепе есть специальное отверстие для непарного глаза, и в случае ископаемых видов это единственный источник информации о нём. Среди современных животных оно встречается только у некоторых рыб [24] [] и рептилий у бесхвостых земноводных бывает только узкий проход для нерва и кровеносного сосуда [80] , а у миног — несквозное углубление в хряще [13].
У костных рыб и четвероногих это отверстие может находиться между теменными или лобными костями. Независимо от этого его называют теменным отверстием [] [] [18] [] или пинеальным отверстием [] лат. Непарный глаз может лежать в самом этом отверстии у большинства рептилий , над черепом у бесхвостых земноводных и некоторых рыб [22] или под ним у миног и некоторых рептилий [79] [80] []. У высших позвоночных теменное отверстие обычно сдвинуто назад по сравнению с его положением у низших, причём тенденция к этому сдвигу заметна уже в ходе эволюции кистепёрых рыб [].
Для рыб более характерно его расположение между лобными костями, а для четвероногих — между теменными [18] [] []. У некоторых звероподобных рептилий например, дейтерозавра оно находилось уже на самом краю затылка []. Это перемещение пинеального отверстия в ходе эволюции объясняется согласно И. Шмальгаузену тем, что промежуточный мозг вместе со своими производными отодвигался назад из-за увеличения конечного мозга [18].
С другой стороны, вопрос о том, какие кости рыб гомологичны теменным и лобным костям четвероногих , очень запутан [18] [] []. Иногда кости, которые окружают пинеальное отверстие у лопастепёрых рыб , считают лобными по определению [] , иногда называют теменными [] , а иногда считают его положение определяющим признаком для теменных костей []. Пинеальное отверстие бывает и на стыке лобных и теменных костей — например, у большинства игуанообразных ящериц, причём у других ящериц этого же инфраотряда оно может располагаться и между теменными, и между лобными, а может и отсутствовать [] [] [] [].
Такие отличия бывают даже между разными особями одного вида [17]. У некоторых животных теменные или лобные кости изначально парные срастаются и пинеальное отверстие оказывается посреди образовавшейся цельной кости. Это наблюдается, например, у вымершей кистепёрой рыбы Osteolepis [] [32] и многих современных ящериц в частности, варанов.
У бесхвостых земноводных непарный глаз лежит над передним краем стыка лобнотеменных костей, которые образуются в результате срастания с каждой стороны лобной кости с теменной [].
У некоторых палеозойских животных — остракодерм , плакодерм и акантод [] [] — деление черепа или головного щита на костные пластины не имело ничего общего с таковым четвероногих, а у некоторых остракодерм вообще отсутствовало. Но их пинеальное отверстие находилось на своём обычном месте — в плоскости симметрии головы поблизости от глазниц между ними или немного спереди [] []. У многих плакодерм [] и остракодерм [] [] оно было расположено на особой маленькой костной пластинке пинеальной пластинке.
У костнопанцирных она была со всех сторон окружена цельным головным щитом, но при этом, вероятно, могла в какой-то мере двигаться вперёд и назад [66]. Для древних кистепёрых характерны 5—6 иногда больше или меньше [] мелких пластинок, окружающих пинеальное отверстие.
Они могли сливаться между собой и с обрамляющими их большими костями []. У некоторых остракодерм вместо сквозного теменного отверстия был утончённый участок кости, известный как пинеальное пятно [] лат.
Панцирь в этом месте мог образовывать пинеальный бугорок []. Приподнятость пинеального отверстия на специальном теменном бугре была характерна и для примитивных терапсид — предков млекопитающих [] [] []. В частности, у титанозуха высота этого бугра могла превышать сантиметр []. Он мог быть образован и только теменными костями, и теменными вместе с лобными []. Какой из двух органов пинеального комплекса содержало это отверстие, для многих ископаемых животных неясно.
Существует мнение, что если оно было расположено спереди например, между передними краями лобных костей , в нём находился пинеальный орган, а если сзади за стыком лобных и теменных — парапинеальный []. Среди вымерших животных были и такие, у которых это отверстие было парным. По некоторым интерпретациям, парное пинеальное [] [42] вероятно, пинеальное с парапинеальным [] [] [] отверстие имели древнейшие из хорошо известных позвоночных с минерализированным скелетом [] — арандаспиды , но по другой версии эти отверстия у них не имели отношения к теменным глазам [].
У некоторых панцирных и костных рыб парность или сдвоенность теменного отверстия установлена точно. Среди панцирных рыб этот признак имели некоторые артродиры Coccosteus [] , Pholidosteus , Rhinosteus [] [16] , Dunkleosteus [] , Titanichthys [] [] и стегоселахии [] [] , а среди костных — некоторые палеониски [] , поролепообразные [] [18] и первые двоякодышащие Dipnorhynchus [en] [] [] [] [] []. Одно из них, а иногда и оба, могло быть не сквозным у палеонисков и поролепообразных.
Кроме того, иногда они были не полностью разделены у некоторых панцирных и двоякодышащих рыб [] [] [] []. Парность этого отверстия у артродир могла варьировать в пределах рода []. Теменное отверстие всегда меньше парных глазниц. У большинства современных рептилий оно меньше миллиметра. Многие ископаемые животные имели гораздо большее [].
Если оно вытянуто, то обычно вдоль оси тела []. У некоторых аделоспондилов Adelogyrinus это отверстие вытягивалось в продольную щель, близкую по длине к глазницам []. Для оценки относительного развития этого отверстия его сравнивают с большим затылочным отверстием лат. У современных ящериц теменное отверстие меньше затылочного как минимум в пять раз, у гаттерии по среднему диаметру — в 3 раза [] , у многих ископаемых амниот они были примерно равны, а у некоторых например, у парейазавра [] и ряда примитивных терапсид [] теменное было даже больше [].
Связь размера теменного глаза с размером теменного отверстия исследована на современных ящерицах. По одним данным, эта связь довольно тесная [] [] [] , а по другим — практически отсутствует [79]. По-видимому, слабая связь этих величин характерна для молодых особей с плохо окостеневшим черепом [] : у них кость охватывает теменной глаз неплотно.
Но он может быть существенно меньше теменного отверстия и у взрослых животных, если окружён толстой соединительнотканной капсулой как у гаттерии []. Теменной глаз явно приспособлен для восприятия света, но во многом его функции до сих пор неясны [2].
Известно, что у миног непарные глаза обеспечивают изменение концентрации мелатонина в организме со временем суток [31] [41] , но многое об их работе ещё предстоит узнать [41]. У ящериц выработка мелатонина теменным глазом невелика у них она происходит в основном в неглазоподобном пинеальном органе [82] [6] [] , и его функции ещё менее понятны. Попытки их определить часто дают неоднозначные, неясные или плохо воспроизводимые результаты.
Большинство исследователей сходится на следующих объяснениях [2] [82] :. У всех современных животных третий глаз, по-видимому, не может давать изображение: это видно и по его строению, и по обычно плохой прозрачности кожи над ним [] [15] [80] []. Эксперименты по поиску поведенческих реакций на его освещение дают противоречивые результаты [] [].
Но у некоторых животных он выполняет не только эндокринные, но и зрительные функции [] : способен чувствовать уровень освещённости, а иногда и направление света и положение плоскости его поляризации [6] [7] [8].
Влияние освещённости на электрическую активность органов пинеального комплекса обнаружено у всех исследованных на этот счёт бесчелюстных, рыб, земноводных и рептилий [29]. В теменном глазу ящериц, в отличие от их пинеального органа, пинеалоциты специализированы для фоторецепции, а не для выработки гормонов [].
Восприятие света с помощью непарных глаз обнаружено у ряда видов из разных классов. У личинок миног это главные светочувствительные органы, поскольку их парные глаза ещё плохо развиты [41] [6]. У некоторых рыб способность чувствовать освещённость пинеальным органом установлена [] , а у некоторых — предполагается [].
Поведенческая реакция на его освещение обнаружена у нерки []. Непарный глаз сомообразных , возможно, реагирует на тени проплывающих сверху объектов [22] []. Некоторым окунеобразным он предположительно помогает ориентироваться при миграциях [] [31]. Светящиеся анчоусы , возможно, используют пинеальный орган вместе с парными глазами для оценки освещённости, что нужно для адекватной контриллюминации — свечения нижней стороны тела, чтобы рыба была менее заметна снизу [].
Есть и данные о визуальной функции непарного глаза лягушек в первую очередь их головастиков : чувствуя этим глазом скачки освещённости, животные начинают двигаться [] [31]. Непарные глаза земноводных и рептилий важны для контроля количества времени, проводимого животным на ярком свету.
Ящерицы и лягушки с искусственно закрытым или удалённым третьим глазом, как правило, дольше греются на солнцепёке что может приводить к перегреву и потерям воды в опасных количествах. Но нарушения их терморегуляции сильно отличаются в зависимости от вида, а иногда и от времени года и суток [82]. Кроме того, у них увеличивается двигательная активность и могут нарушаться суточные ритмы [82] [27] [10] [4] [5].
Третий глаз земноводных и рептилий, в отличие от парных глаз [] , чувствителен к положению плоскости поляризации света это обеспечивается тем, что мембранные диски в пинеалоцитах, расположенных по краям его сетчатки, параллельны падающему свету [8] [] и некоторые животные используют это его свойство для ориентировки в пространстве [7] [] [6] [] [82].
Это возможно благодаря тому, что чистое небо даёт довольно сильно поляризованный свет. Эксперименты показывают, что восприятие света теменным глазом необходимо для ориентировки таким способом у руинной ящерицы [7].
Эта функция установлена даже для пинеального органа тигровой амбистомы , мало похожего на глаз [8] [6] [] []. Показано, что у некоторых животных сверчковой квакши и лягушки-быка [6] третий глаз участвует в ориентировке по азимуту Солнца.
Необходимость его работы для ориентировки в пространстве обнаружена и ещё у некоторых видов короткохвостого сцинка [] , заборной игуаны [7] , песчаной игуаны [] [] , но её конкретный механизм у них не выяснен. Возможно, изменения поведения ящериц с закрытым теменным глазом в частности, увеличение активности связаны именно с нарушением ориентации []. С другой стороны, есть версия, что с помощью теменного глаза ящерицы ориентируются не столько по Солнцу или поляризованному свету, сколько по магнитному полю Земли [9].
Это мнение основывается на том, что у многих животных обнаружена необходимость освещения органов, которые обеспечивают магниторецепцию, для их работы [9] [] , причём у тритонов [] [] [6] [19] [9] и, предположительно, у птиц [19] [] эта функция локализована в пинеальном органе.
Амфибии способны с его помощью ориентироваться и по магнитному полю, и по положению плоскости поляризации света [] ; возможно, восприятие этих факторов обеспечивает один и тот же биофизический механизм []. Третий глаз земноводных и рептилий реагирует на излучение в широком диапазоне длин волн: от ультрафиолетового до красного [8] [31].
Максимум чувствительности пинеального органа миног [] [] [31] , рыб [] и лягушек [] и парапинеального органа ящериц [6] [31] лежит в зелёной или синей области. К инфракрасному излучению а также к термическим, механическим и химическим воздействиям непарные глаза нечувствительны [8]. Это исключает предполагавшуюся для них ранее [] терморецепторную функцию [82]. У лягушек третий глаз почти не адаптируется к изменениям освещённости что даёт возможность определять её интенсивность [4].
У ящериц эта адаптация выражена намного сильнее — даже лучше, чем у их парных глаз: например, чувствительность теменного глаза зелёной игуаны после часа пребывания в темноте увеличивается почти на два порядка у парных глаз — на один [98]. У лягушек минимальный интервал между стимулами, которые этот глаз способен чувствовать раздельно, составляет около 2 минут, а у ящериц — десятые доли секунды [4] [98] , что на порядок больше, чем у их парных глаз [98].
Пинеалоциты, в отличие от всех остальных известных фоторецепторов , совмещают два противоположных способа реакции на свет. Один из их опсинов с максимумом чувствительности в зелёной области при поглощении света вызывает деполяризацию клетки, а второй с максимумом чувствительности в синей области — её гиперполяризацию. Эти опсины связаны с разными биохимическими путями передачи сигнала [] [] [].
У колбочек и палочек парных глаз при освещении наблюдается только гиперполяризация []. При постоянном освещении частота импульсов в нерве теменного глаза уменьшается пропорционально логарифму интенсивности света. При освещении короткими вспышками разные ганглионарные клетки проявляют разную зависимость электрической активности от интенсивности и длины волны света. Они делятся на два типа: хроматические и ахроматические [29] [].
Ахроматические клетки уменьшают свою активность в ответ на освещение любого цвета. Активность хроматических клеток подавляется синим светом и усиливается более длинноволновым зелёным — красным [] [4]. Миелинизированы только волокна, идущие от ахроматических клеток [8]. Своеобразное поведение хроматических клеток, вероятно, вызвано способностью пинеалоцитов реагировать на свет разного цвета противоположными способами [] []. Основная эндокринная функция непарных глаз, как и других органов пинеального комплекса — синтез мелатонина , происходящий в пинеалоцитах.
Небольшое количество этого гормона образуется и в сетчатке парных глаз [20] [] [3] , а также в некоторых других органах, но только в пинеальном комплексе и иногда [20] сетчатке парных глаз интенсивность его синтеза зависит от времени суток — почти у всех видов она ночью увеличивается [] [3]. Кроме того, в пинеальном комплексе синтезируются другие индоламины в том числе серотонин , из которого и образуется мелатонин [] и ряд пептидов [] [20].
У миног непарные глаза вырабатывают существенное количество названных веществ [31] [41]. У ящериц доля глазоподобного органа в общей их выработке пинеальным комплексом невелика возможно, он синтезирует мелатонин только для своих собственных нужд [6] [] [82].

Невелико и его влияние на эндокринную активность пинеального органа [82]. Тем не менее он и у ящериц [82] [5] [27] , и у лягушек [6] вносит вместе с парными глазами и фоторецепторами мозга некоторый вклад в синхронизацию суточных ритмов организма с циклом смены дня и ночи, а также участвует в регуляции размножения [82] [6]. У животных, сохранивших теменной глаз или светочувствительность пинеального органа то есть у большинства позвоночных эндокринную активность пинеального комплекса регулирует освещение.
У тех животных, которые потеряли светочувствительность этих органов у млекопитающих и, в некоторой степени, птиц активность пинеального органа регулирует мозг а именно супрахиазматическое ядро в гипоталамусе , которое у этих животных служит главными биологическими часами. Ритм активности этого ядра, в свою очередь, корректируется информацией, которая поступает из сетчатки глаз [] [3] [31].
Но есть данные, что у новорождённых крыс пинеальный орган способен непосредственно реагировать на свет, причём он его получает больше, чем глаза ещё не открытые []. Кроме того, у многих видов непарный глаз как и другие органы пинеального комплекса, а также сетчатка парных глаз работает как биологические часы : способен к поддержанию суточного ритма синтеза мелатонина даже без внешних световых и нервных стимулов [] [3] [6] [94].
Мелатонин участвует в регуляции многих процессов с суточной или сезонной ритмичностью, в частности, сна, зимней спячки , колебаний температуры тела, двигательной и кормовой активности, миграции, линьки, размножения и изменений окраски [] [36] [].
У большинства видов мелатонин даёт антигонадотропный эффект, у некоторых — противоположный [20] [59]. Подобным действием обладают и некоторые другие индолеамины и полипептиды, синтезирующиеся в пинеальном комплексе []. У миног эндокринное влияние непарного глаза вызывает рост двигательной активности с наступлением темноты [31] []. Установлено, что у взрослых миног у которых исчезает суточная ритмичность поведения пинеальный комплекс перестаёт вырабатывать мелатонин, хотя в сетчатке парных глаз его синтез продолжается [].
У некоторых рыб [] [14] и головастиков лягушек [6] он управляет и суточными вертикальными миграциями. У ящериц теменной глаз подавляет двигательную активность [10] [27] , но её суточные изменения, видимо, не зависят от его влияния [6] [82].
Теменной глаз многих животных управляет суточными изменениями окраски: под его влиянием кожа светлеет ночью и темнеет днём. Это известно у миног [] , некоторых рыб [] , земноводных и рептилий [] [20] [59] [82]. Пинеальный комплекс, в том числе, возможно, теменной глаз ящериц [6] [82] , участвует и в терморегуляции [4] , но в механизме этой связи остаётся много неясного [2] [].
С его терморегуляторной функцией связывают то, что он чаще всего лучше развит у жителей высоких широт [60] [2]. Таким образом, концентрация мелатонина служит показателем времени суток, и от неё зависит множество разных циклических процессов [3] , причём её влияние на эти процессы у разных видов может быть разным [].
Кроме того, есть данные, что мелатонин участвует в адаптации сетчатки к изменениям освещённости и служит антиоксидантом [] [3]. Развитие у предков позвоночных теменного глаза, возможно, объясняется их придонным образом жизни. Способность видеть угрозу сверху для таких животных очень важна.
Для их потомков, которые перешли к активной жизни в толще воды, возможность смотреть вверх потеряла актуальность [12]. В таком случае теменной глаз современных позвоночных — рудиментарный орган. В пользу этого говорит и его потеря многими группами независимо друг от друга [2] , и то, что он напоминает по строению редуцированные парные глаза для которых характерно погружение под кожу и потеря преломляющего аппарата и нерва []. Но в некоторых эволюционных линиях его зрительная функция могла сохраняться ещё долго у некоторых форм в некоторой мере, видимо, и до сих пор.
В частности, на это указывает размер теменного отверстия многих вымерших рептилий [] [80] и данные по некоторым современным животным [] [44] [].
С другой стороны, возможно, что органы пинеального комплекса никогда не были развиты настолько, чтобы делать большой вклад в зрение животного. Не исключено, что эндокринная функция изначально была для них главной [3] [34] , а глазоподобное строение этих органов в отдельных эволюционных линиях развилось позже [15] [].
В пользу того, что эти два органа изначально были правым и левым в паре, говорит характер их нервных соединений [31] [27] , а также парность или сдвоенность теменного отверстия или несквозных углублений в черепе у некоторых ископаемых животных [16] [] : арандаспид возможно [42] [] , некоторых панцирных рыб , палеонисков и лопастепёрых [] [] [] [16] [18].
По другой версии и парапинеальный, и, возможно, пинеальный орган изначально были парными [16] [] [33] [15] , то есть у предков позвоночных было не меньше 5—6 светочувствительных органов. Согласно этой гипотезе, правый парапинеальный орган в ходе эволюции исчез или вошёл в состав левого , а правый и левый пинеальные слились [16].
На это указывают, в частности, наблюдения случаев парности парапинеального органа у миног [] и ящериц [62] [15] [79] [39]. В пользу этой версии говорят и некоторые эмбриологические данные, в том числе результаты исследования экспрессии генов [68] [16]. Существует гипотеза, что у предков позвоночных было несколько гомологичных пар органов зрения, и от одной из них произошли парные глаза современных форм, а от другой расположенной позади — их пинеальный и парапинеальный орган [26].
На это указывают эмбриологические исследования акул [] [] [] [] []. Среди беспозвоночных гомолог пинеального комплекса найден у ланцетника. Это так называемое ламеллярное тело англ. Это установлено по микроструктуре и гистохимии этих органов [] , а также по исследованию экспрессии генов в них []. Они не имеют глазоподобной формы, но содержат ресничные фоторецепторы []. В XIX и начале XX века были популярны предположения и утверждения [54] о гомологии теменного глаза позвоночных медианному глазу оболочников и даже некоторых членистоногих , основанные на расположении этих глаз и неинвертированности их сетчатки [] [] [] [] [].
В эволюции позвоночных видна тенденция к редукции этого органа [2].

Теменное отверстие имели многие палеозойские животные, но начиная с триаса оно стало встречаться существенно реже [12] [80]. По систематическому положению животных, имеющих теменной глаз, видно, что он независимо исчезал во многих группах [2]. Потерю этого органа предками млекопитающих и птиц объясняют по-разному. Согласно одной версии, это следствие развития у них гомойотермии.
По-видимому, одна из функций теменного глаза рептилий — участие в терморегуляции в качестве датчика интенсивности солнечного света. С появлением эффективных механизмов терморегуляции эта функция потеряла актуальность [] [] [10].
По другой версии предки млекопитающих утратили теменной глаз из-за своего ночного образа жизни []. Но и млекопитающие, и птицы сохранили пинеальный орган, который работает как железа внутренней секреции как и у остальных позвоночных. На этом основано мнение, что в ходе эволюции органов пинеального комплекса менялись не столько их функции, сколько способ получения ими информации об освещённости с помощью собственной сетчатки у низших позвоночных и через мозг у высших [14].
Соответственно, постепенно развивалась эфферентная иннервация этих органов и терялась афферентная []. Такие изменения видны и в онтогенезе млекопитающих: эфферентная иннервация их пинеального органа появляется тогда, когда исчезают его светочувствительные элементы [].
Фоторецепторные клетки пинеального комплекса пинеалоциты [en] и парных глаз палочки и колбочки , по всей видимости, имеют общее происхождение — они развились из клеток предков позвоночных, которые были способны и к фоторецепции, и к синтезу мелатонина. В ходе эволюции между ними появилось разделение функций, лучше всего выраженное у млекопитающих и, в меньшей степени, у птиц : фоторецепторы парных глаз обеспечивают зрение, а пинеалоциты специализируются на выработке мелатонина — гормона , регулирующего различные биоритмы [3].
Тем не менее небольшое количество мелатонина образуется и в сетчатке парных глаз. Там он является продуктом реакций детоксикации вредных для сетчатки соединений — арилалкиламинов , в частности, серотонина они реагируют с необходимым для зрения ретинальдегидом , причём в результате образуются ядовитые вещества [3] [].
Кроме того, мелатонин выполняет в сетчатке некоторые функции [] [3] — повышает её светочувствительность путём перераспределения меланосом в её пигментных клетках аналогично он влияет и на меланоциты кожи [20] и, возможно, служит антиоксидантом [].
Вероятно, у предков позвоночных как и у современных низших хордовых мелатонин не был гормоном. Он был отходом метаболизма сетчатки [] [3] или выполнял функции, нужные только в ней [20] [].
Одной из адаптаций сетчатки к суточным изменениям освещённости стало повышение ночью интенсивности реакций, приводящих к синтезу мелатонина что могло быть связано с потребностью в нём самом или просто в детоксикации серотонина.
Вследствие этого появились цикличные изменения концентрации мелатонина, и она стала фактически показателем времени суток. Тогда развилась зависимость от неё различных циклических процессов: мелатонин стал регулятором суточных ритмов [] [3] [20]. В ходе развития и усиления этой зависимости увеличилась и потребность в мелатонине: его понадобилось больше, чем образовывалось в ходе нормального функционирования сетчатки.
Но повышение его выработки было несовместимым со зрительной функцией глаз, так как оно требует увеличения количества в сетчатке серотонина, который для неё вреден.
Вероятно, это и привело к тому, что эндокринная и зрительная функция разделились между разными органами [] [3] []. Распространённое мнение, что хорошо развитый теменной глаз бывает только у относительно примитивных форм, не подтверждается детальными исследованиями, охватывающими большое число ископаемых и современных позвоночных [] [] , хотя тенденция к его исчезновению видна в эволюции многих групп [2] [] [] [] [].
Не видно по этим исследованиям и тесной связи степени развития этого органа с образом жизни животного [] [11] [2] , хотя некоторые закономерности здесь прослеживаются. Замечена тенденция к лучшему развитию теменного глаза и других органов пинеального комплекса у животных с интенсивным метаболизмом и у жителей более высоких широт, что связывают с его терморегуляторной функцией [60].
Но есть и много исключений из этих закономерностей. Так, более сильное развитие этих органов у видов высоких широт прослеживается у ящериц [60] [82] и млекопитающих [] [2] , но не у земноводных [60] []. Исследование тридцати видов ящериц рода Liolaemus показало, что размер их теменного глаза при своей большой изменчивости не зависит от климатических условий [2].
По вымершим рептилиям замечено, что большое теменное отверстие более характерно для больших медлительных травоядных например, диадектов , казеид , больших проколофономорф , хотя встречается и у мелких хищников [].
Плохая развитость теменного глаза характерна для древесных ящериц. В частности, у большинства хамелеонов он сильно упрощён, а у остальных отсутствует. В тех семействах, у большинства представителей которых этот орган развит хорошо, исключениями часто являются именно формы с древесным образом жизни [75]. У ящериц, ведущих подземный образ жизни, он обычно не исчезает кроме тех, у которых редуцированы и парные глаза [75].
Органы пинеального комплекса, в том числе теменной глаз, обычно лучше развиты у дневных животных, чем у ночных. Это прослеживается и у рептилий, и у птиц, и у млекопитающих, хотя во всех этих классах есть и исключения [60] [] [].
Тот факт, что млекопитающие практически потеряли светочувствительность пинеального органа, иногда связывают с тем, что ранние представители этого класса были ночными животными. Миксины потеряли и сам пинеальный комплекс, и это, возможно, следствие того, что они проводят большую часть жизни в тёмных придонных местообитаниях [] []. Но среди рыб пинеальное окно [20] [31] и хорошо развитые пинеалоциты [] [] встречаются чаще у глубоководных видов, чем у мелководных, хотя у тех, которые живут в полной темноте — на глубинах больше м и в пещерах — этого окна не бывает как и отверстия в черепе.
Но пинеальный орган, в отличие от парных глаз, у них обычно не редуцируется [] [] []. Остракодермы — покрытые панцирем палеозойские бесчелюстные — обычно имели в головном костном щите отверстие для третьего глаза. Оно было расположено примерно между парными глазницами позади ноздрей [] []. Пинеальное отверстие у остракодерм часто было довольно крупным, но как и у других животных никогда не больше парных глазниц [12]. Такое отверстие было уже у древнейших хорошо изученных остракодерм [] — арандаспид , причём у них оно было парным [] [42].
По некоторым интерпретациям, это были отверстия для пинеального и парапинеального органа [] [] [] , но по некоторым — они не имели к ним никакого отношения и служили выходами эндолимфатических протоков []. У всех остальных остракодерм было не больше одного пинеального отверстия. Оно характерно для костнопанцирных [] , беспанцирных [] и примитивных представителей разнощитковых и галеаспид [66].
Миноги кроме Mordacia имеют, помимо парных глаз , два непарных: пинеальный лучше развит и парапинеальный.
Южная минога Mordacia имеет только пинеальный орган, причём плохо развитый [16] []. Теменного отверстия у миног нет, но крыша черепа над пинеальным комплексом утоньшается [13] [] [63] [32]. Есть пинеальное окно — непигментированный участок кожи позади единственной ноздри [90] [] []. У миксин пинеальный комплекс отсутствует начисто [] [15] , а управление суточными циклами обеспечивают рудиментарные парные глаза [].
Не найдены у миксин и рецепторы мелатонина, и на этом основано предположение, что их предки отделились от предков остальных позвоночных ещё до появления у тех регулировки биоритмов с его помощью []. По одной из версий, пинеальный комплекс миксин редуцировался в ходе эволюции [] [94] [19] [].
С другой стороны, есть гипотеза, что от него и произошли их парные глаза, а у остальных позвоночных они возникли независимо [].
Глаза миксин во многих отношениях больше похожи на непарные глаза других животных, чем на парные: у них нет радужки, роговицы и хрусталика, а их светочувствительные клетки соединяются с ганглионарными напрямую [34] [] [31].
Но в глазах миксин, в отличие от непарных глаз, сетчатка инвертирована []. Есть и сообщения о наблюдениях у миксины зачатка пинеального органа на ранних стадиях развития [] , а также атавистического непарного глаза у взрослой особи []. У многих панцирных рыб силур — девон третий глаз был хорошо развит, причём у некоторых например, Pholidosteus , Rhinosteus [16] , Titanichthys [] , Coccosteus соответствующее отверстие в черепе иногда было парным или сдвоенным [].
Эти данные говорят в пользу версии о происхождении пинеального комплекса от пары правого и левого органов [16]. Находился он у них немного спереди от парных глаз [] []. Небольшой третий глаз был, в частности, у гигантской панцирной рыбы дунклеостеуса [] [].
У Acanthodes — единственного хорошо изученного рода акантод — было отверстие в черепе, которое, вероятно, было пинеальным [] [].
У хрящевых рыб парапинеальный орган не известен [20] [19] , а пинеальный есть почти у всех представителей но есть данные, что он образуется из двух сливающихся пузырьков [61]. Он имеет вид длинной трубки [67] со вздутиями на концах есть предположение, что пинеальному и парапинеальному органам других животных соответствуют эти вздутия [31] []. Его конец лежит близко к поверхности кожи [].
Но у электрических скатов из рода Torpedo пинеального комплекса нет вообще [59] [94] []. Пинеальное окно есть у многих пелагических акул [] , а у некоторых кархаринообразных [44] [] и катранообразных [] [] [] есть даже отверстие в черепе [] [14] [15] [94] []. Но, вообще говоря, для акул и современных, и вымерших пинеальное отверстие — редкость [12]. У костных рыб обычно хорошо развит пинеальный орган и плохо — парапинеальный [38] [] [94].
Тем не менее он есть у некоторых видов и лопастепёрых , и лучепёрых рыб. Череп над этими органами часто тоньше, чем в других местах [] , но отверстия в нём у большинства современных костных рыб, как и у большинства хрящевых, нет [16] [95].
К исключениям относятся, например, веслонос , многие сомообразные [] например, панцирные и угрехвостые сомы [24] , Loricaria [] [33] , Sciades [22] , карпообразные Labeo [38] , окунеобразные тунец , меч-рыба и другие []. У палеозойских особенно девонских костных рыб это отверстие встречается намного чаще. Особенно характерно оно для кистепёрых [12] , хотя у представителей современного их рода — латимерии — его нет []. У лучепёрых , как и у других рыб, пинеальный орган обычно развит.
Особенно это характерно для умеренно глубоководных и для совершающих вертикальные миграции видов. Чтобы ее переработать систематизировать, соотнести с уже имеющейся информацией, обобщить и уложить в памяти , необходимы время и подходящие условия. Получив дозу сведений за день, мозг подводит итоги. В бодрствующем состоянии мысль остается в рамках шаблонов.
Во сне иная ситуация. И если есть необходимые знания и информация, но не хватает лишь способа по-особому, необычно их соединить, то состояние сна — самое подходящее время для прозрения.
При методическом обдумывании оно не наступит. День уступает свои права ночи, и вот, наконец, мы чувствуем сонливость. Состояние сонливости — это результат подготовки мозга к ночному режиму, режиму сна.
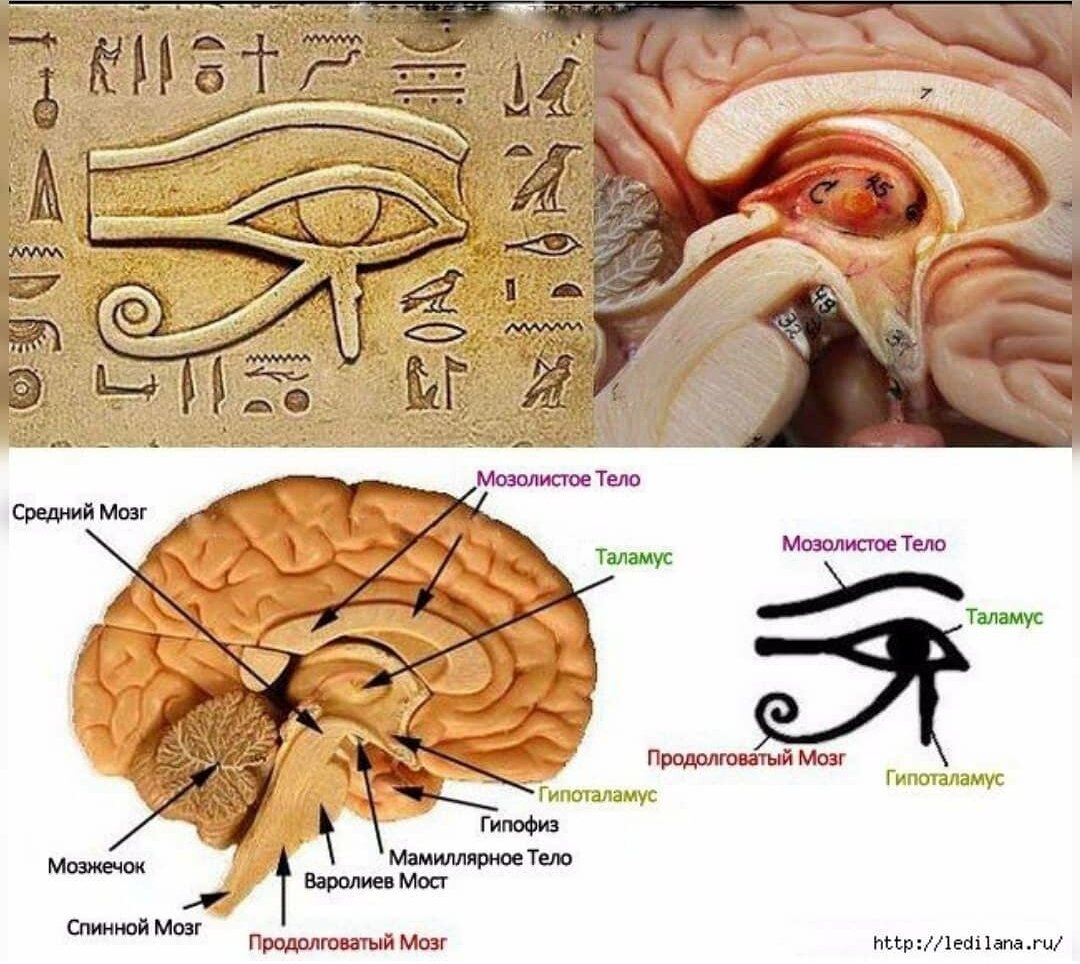
Почему во сне решения приходят быстро, а мысли точны и даже гениальны? Мысль — это своеобразное «семя», и для того, чтобы оно созрело и выросло, нам необходимо посадить его, то есть поместить в темноту. Мелатонин и сератонин производятся в основном в головном мозге шишковидной железой. Она расположена в центре мозга и связана с глазами двумя «канатиками». Другими словами, в шишковидной железе и в глазах содержатся одинаковые клетки. Многие ученые сделали заключение, что шишковидная железа — это и есть «третий глаз».
В этом есть правда. Шишковидная железа оценивает свет и темноту, и в тот момент, когда глаза закрываются и перестают видеть свет, она вместо сератонина начинает выделять мелатонин.
Мелатонин замедляет Ваши процессы и успокаивает Вас. Он — предшественник пинолина, который выделяется во время глубокого сна.
Замедлив Вас, он позволит Вам попасть в состояние осознанного сна. Вот почему так важно ложиться спать с чистыми мыслями и позитивным настроем. Мозг воспринимает все буквально!
Иными словами, если Вы ложитесь спать уставшими и раздражительными, утро окажется далеко «не добрым». Большинство из нас надеется убежать от тягот жизни. Однако сон — это шанс вернуться к радости своего истинного предназначения», — Генри Рид. Какой сон выбрать — зависит от каждого из нас. Выспимся ли мы сегодня или бодрость по утрам останется только призрачной мечтой?
Жизнь многогранна, хочется успеть многое, но хватит ли сил? Где искать резервы здоровья и настроения? Как известно, резерв есть всегда, и мы предлагаем его — это тот запас отдыха и здоровья, который Вы можете приобрести вместе с новым удобным анатомическим матрасом. Главной движущей силой компании «Аскона» в создании товаров для сна является осознание важности процесса сна и его роли в жизни человека.
Специалисты компании, профессионалы производства матрасов, знают, как сделать матрас с заданными характеристиками, используя современные составляющие. Создать условия для здорового сна невозможно без оценки его физиологических аспектов.
Вследствие земного притяжения участки нашего тела, соприкасающиеся с поверхностью, на которой мы спим, испытывают давление. Для разгрузки этих участков мы периодически переворачиваемся во сне. При избыточном воздействии давления на мягкие ткани и кожные покровы происходит прекращение нормального кровоснабжения сдавленных участков. Равномерное распределение веса тела по поверхности, на которой мы спим, снижает общее давление на мягкие ткани и влечет за собой более спокойный сон.
Были проведены исследования физиологической реакции тела человека на твердость поверхности, на которой он спит. Это показывает, насколько велико влияние характеристик спальной поверхности на качество сна. Таким образом, хороший матрас для сна с оптимальными поддерживающими характеристиками вместе с правильно подобранной анатомической подушкой способны сделать Ваш сон глубже, тем самым увеличивая возможность появления «озарений» и гениальных мыслей.