Основными источниками энергии для организма являются
В результате отмечается появление сухости и шелушения кожи, трещинки на губах, угревая сыпь; ломкость и выпадение волос, их истончение и потускнение цвета; ломкость ногтей, утрата их глянцевого слоя. Textbook of Medical Physiology неопр. Широкое использование пищевых добавок производителями пищевых продуктов определяется стремлением - увеличить срок сохранения качества пищевых продуктов, в том числе скоропортящихся и быстро черствеющих в условиях необходимости их перевозки на большие расстояния; удовлетворить потребителя высокими и насыщенными вкусовыми качествами продукта связано с использованием ароматизаторов и красителей; создание новых видов пищевых продуктов, отвечающих современным требованиям науки о питании низкокалорийные продукты, имитаторы продуктов - это связано с использованием пищевых добавок, регулирующих консистенцию пищевых продуктов. У здорового человека в течение суток количество распадающегося белка должно соответствовать количеству вновь синтезированного.
Энергия ветра обусловлена неравномерностью прогрева земной поверхности, энергия углеводородов рождается под влиянием фотосинтеза, гидроэнергия рек образуется от испарения воды и последующего выпадения осадков. Только несколько видов альтернативных источников не имеют солнечную природу. Это энергия приливов и отливов, обусловленная гравитационным притяжением Луны, ядерная энергия, запасенная вселенной много миллиардов лет назад и геотермальная энергия Земли, образованная при ее формировании.
Эти виды энергии составляют незначительную долю в энергетическом балансе планеты. Можно сказать, что Солнце — это единственный основной источник энергии и жизни на Земле. При этом выделяется гигантское количество энергии, которая распространяется в виде радиации и доходит до Земли. Здесь и происходит ее преобразование в другие виды энергии. Солнечную энергию мы можем превратить, например, в электрическую, используя эффект фотосинтеза.
Солнце в масштабах существования человеческой цивилизации является неисчерпаемым источником энергии. Альтернативная энергетика как раз и использует преобразованную солнечную энергию. Главное преимущество ее в том, что в основном — это экологически чистые источники энергии. Традиционная энергетика исторически сопровождалась выбросами вредных веществ, превышением нормы углекислого газа в атмосфере, что приводило к парниковому эффекту и глобальному потеплению.
Солнечная радиация напрямую превращается в электричество, ветровые установки тоже не несут выбросов. Но у альтернативных источников есть один существенный недостаток.
Это — нестабильность генерации энергии, которая зависит от природных условий. Промышленность и крупные мегаполисы не могут полностью полагаться на такой ненадежный источник. Альтернативная энергетика получит мощный импульс развития, если решит проблему по накоплению энергии, причем объемы накопления должны быть гигантские, соизмеримые с существующими запасами углеводородного топлива.
Создание такой технологии накопления электричества названо McKinsey Global Institute одной из 12 прорывных технологий, которые существенным образом изменят глобальную экономику.
На Энергетическом факультете ведутся такие работы на базе водородной энергетики. У нас хорошо развито изучение традиционной энергии: это электрические станции, сети, системы электроснабжения. Имеется направление магистратуры, на котором мы готовим специалистов по альтернативной энергетике. Также у нас имеются серьезные научные заделы в этом направлении, в том числе выполненные совместно с американскими учеными. При этом энергия от окисления этих соединений образуется с помощью акцепторов электронов, таких как кислород или нитриты [51].
Процессы получения энергии из неорганических веществ играют важную роль в таких биогеохимических циклах, как ацетогенез , нитрификация и денитрификация [52] [53]. Энергия солнечного света поглощается растениями , цианобактериями , пурпурными бактериями , зелёными серными бактериями и некоторыми простейшими. Этот процесс часто сочетается с превращением диоксида углерода в органические соединения, как часть процесса фотосинтеза см. Системы захвата энергии и фиксации углерода у некоторых прокариот могут работать раздельно например у пурпурных и зелёных серных бактерий [54] [55].
У многих организмов поглощение солнечной энергии в принципе аналогично окислительному фосфорилированию, так как при этом энергия запасается в форме градиента концентрации протонов и движущая сила протонов приводит к синтезу АТФ [17].
Электроны, необходимые для этой цепи переноса, поступают от светособирающих белков, называемых центрами фотосинтетических реакций примером являются родопсины. В зависимости от вида фотосинтетических пигментов классифицируют два типа центров реакций; в настоящее время большинство фотосинтезирующих бактерий имеют только один тип, в то время как растения и цианобактерии два [56].
У растений, водорослей и цианобактерий фотосистема II использует энергию света для удаления электронов из воды, при этом молекулярный кислород выделяется как побочный продукт реакции. Электроны затем поступают в комплекс цитохрома b6f, который использует энергию для перекачки протонов через тилакоидную мембрану в хлоропластах [7].
Под действием электрохимического градиента протоны движутся обратно через мембрану и запускают АТР-синтазу.
Анаболизм — совокупность метаболических процессов биосинтеза сложных молекул с затратой энергии. Сложные молекулы, входящие в состав клеточных структур, синтезируются последовательно из более простых предшественников.
Анаболизм включает три основных этапа, каждый из которых катализируется специализированным ферментом. На первом этапе синтезируются молекулы-предшественники, например аминокислоты , моносахариды , терпеноиды и нуклеотиды. На втором этапе предшественники с затратой энергии АТФ преобразуются в активированные формы. На третьем этапе активированные мономеры объединяются в более сложные молекулы, например белки , полисахариды , липиды и нуклеиновые кислоты.
Не все живые организмы могут синтезировать все биологически активные молекулы. Автотрофы например растения могут синтезировать сложные органические молекулы из таких простых неорганических низкомолекулярных веществ, как углекислый газ и вода. Гетеротрофам необходим источник более сложных веществ, таких как моносахариды и аминокислоты, для создания более сложных молекул.
Организмы классифицируют по их основным источникам энергии: фотоавтотрофы и фотогетеротрофы получают энергию из солнечного света, в то время как хемоавтотрофы и хемогетеротрофы получают энергию из неорганических реакций окисления. Фотосинтезом называют процесс биосинтеза сахаров из углекислого газа, при котором необходимая энергия поглощается из солнечного света. У растений , цианобактерий и водорослей при кислородном фотосинтезе происходит фотолиз воды, при этом как побочный продукт выделяется кислород.
Реакция связывания углерода осуществляется с помощью фермента рибулозобисфосфаткарбоксилазы и является частью цикла Кальвина [58].
У растений классифицируют три типа фотосинтеза — по пути трёхуглеродных молекул, по пути четырёхуглеродых молекул С4 , и CAM фотосинтез. Три типа фотосинтеза отличаются по пути связывания углекислого газа и его вхождения в цикл Кальвина; у C3 растений связывание CO 2 происходит непосредственно в цикле Кальвина, а при С4 и CAM CO 2 предварительно включается в состав других соединений. Разные формы фотосинтеза являются приспособлениями к интенсивному потоку солнечных лучей и к сухим условиям [59].
У фотосинтезирующих прокариот механизмы связывания углерода более разнообразны. Углекислый газ может быть фиксирован в цикле Кальвина, в обратном цикле Кребса [60] или в реакциях карбоксилирования ацетил-КоА [61] [62]. Прокариоты — хемоавтотрофы также связывают CO 2 через цикл Кальвина, но для протекания реакции используют энергию из неорганических соединений [63].
В процессе анаболизма сахаров простые органические кислоты могут быть преобразованы в моносахариды , например в глюкозу , и затем использованы для синтеза полисахаридов , таких как крахмал.
Образование глюкозы из соединений, таких как пируват , лактат , глицерин , 3-фосфоглицерат и аминокислот , называют глюконеогенезом.
В процессе глюконеогенеза пируват превращается в глюкозофосфат через ряд промежуточных соединений, многие из которых образуются и при гликолизе [41]. Однако глюконеогенез не просто является гликолизом в обратном направлении, так как несколько химических реакций катализируют специальные ферменты, что даёт возможность независимо регулировать процессы образования и распада глюкозы [64] [65].
Многие организмы запасают питательные вещества в форме липидов и жиров, однако позвоночные не имеют ферментов, катализирующих превращение ацетил-КоА продукта метаболизма жирных кислот в пируват субстрат глюконеогенеза [66]. После длительного голодания позвоночные начинают синтезировать кетоновые тела из жирных кислот, которые могут заменять глюкозу в таких тканях, как головной мозг [67].
У растений и бактерий данная метаболическая проблема решается использованием глиоксилатного цикла , который обходит этап декарбоксилирования в цикле лимонной кислоты и позволяет превращать ацетил-КоА в оксалоацетат и далее использовать для синтеза глюкозы [66] [68]. Помимо жира, глюкоза хранится в большинстве тканей в качестве энергетического ресурса, доступного в тканях посредством гликогенеза, который обычно используется для поддержания уровня глюкозы в крови [69].
Полисахариды и гликаны образуются путем последовательного добавления моносахаридов гликозилтрансферазой из реактивного донора сахара-фосфата, такого как уридиндифосфат глюкозы UDP-Glc , к акцепторной гидроксильной группе на растущем полисахариде.
Поскольку любая из гидроксильных групп в кольце субстрата может быть акцепторами, полученные полисахариды могут иметь прямые или разветвленные структуры [70]. Полисахариды выполняют структурные и метаболические функции, а также могут быть соединены с липидами гликолипиды и белками гликопротеиды при помощи ферментов олигосахаридтрансфераз [71] [72]. Жирные кислоты образуются синтазами жирных кислот из ацетил-КоА.
Углеродный скелет жирных кислот удлиняется в цикле реакций, в которых сначала присоединяется ацетильная группа, далее карбонильная группа восстанавливается до гидроксильной, затем происходит дегидратация и последующее восстановление. Ферменты биосинтеза жирных кислот классифицируют на две группы: у животных и грибов все реакции синтеза жирных кислот осуществляются одним многофункциональным белком I типа [73] , в пластидах растений и у бактерий каждый этап катализируют отдельные ферменты II типа [74] [75].
Терпены и терпеноиды являются представителями самого многочисленного класса растительных натуральных продуктов [76]. Представители данной группы веществ являются производными изопрена и образуются из активированных предшественников изопентилпирофосфата и диметилаллилпирофосфата , которые, в свою очередь, образуются в разных реакциях обмена веществ [77]. У животных и архей изопентилпирофосфат и диметилаллилпирофосфат синтезируются из ацетил-КоА в мевалонатном пути [78] , в то время как у растений и бактерий субстратами не-мевалонатного пути являются пируват и глицеральдегидфосфат [77] [79].
В реакциях биосинтеза стероидов молекулы изопрена объединяются и образуют сквалены , которые далее формируют циклические структуры с образованием ланостерола [80]. Ланостерол может быть преобразован в другие стероиды, например холестерин и эргостерин [80] [81].
Организмы различаются по способности к синтезу 20 общих аминокислот. Большинство бактерий и растений могут синтезировать все 20, но млекопитающие способны синтезировать лишь 10 заменимых аминокислот [7].
Таким образом, в случае млекопитающих 9 незаменимых аминокислот должны быть получены из пищи. Некоторые простые паразиты, такие как бактерии Mycoplasma pneumoniae , не синтезируют все аминокислоты и получают их непосредственно от своих хозяев [82]. Все аминокислоты синтезируются из промежуточных продуктов гликолиза , цикла лимонной кислоты или пентозомонофосфатного пути.
Перенос аминогрупп с аминокислот на альфа-кетокислоты называется трансаминированием. Донорами аминогрупп являются глутамат и глутамин [83]. Второстепенный синтез аминокислот зависит от образования соответствующей альфа-кетокислоты, которая затем подвергается трансаминированию с образованием аминокислоты [84]. Аминокислоты, соединёнными пептидными связями , образуют белки.
Каждый белок имеет уникальную последовательность аминокислотных остатков первичная структура белка. Подобно тому, как буквы алфавита могут комбинироваться с образованием почти бесконечных вариаций слов, аминокислоты могут связываться в той или иной последовательности и формировать разнообразные белки.
Белки состоят из аминокислот, которые были активированы путем присоединения к молекуле переносной РНК через эфирную связь. Аминоацил-тРНК являются субстратами для рибосом , которая объединяют аминокислоты в длинные полипептидные цепочки, используя матрицу мРНК [86]. Нуклеотиды образуются из аминокислот, углекислого газа и муравьиной кислоты в цепи реакций, для протекания которых требуется большое количество энергии [87] [88]. Именно поэтому большинство организмов имеют эффективные системы сохранения ранее синтезированных нуклеотидов и азотистых оснований [87] [89].
Пурины синтезируются как нуклеозиды в основном связанные с рибозой. Аденин и гуанин образуются из инозин -монофосфата, который синтезируется из глицина , глутамина и аспартата при участии метенил- тетрагидрофолата. Пиримидины синтезируются из оротата , который образуется из глутамина и аспартата [90].
Все организмы постоянно подвергаются воздействию соединений, накопление которых может быть вредно для клеток.
Такие потенциально опасные чужеродные соединения называются ксенобиотиками [91]. Ксенобиотики, например синтетические лекарства и яды природного происхождения, детоксифицируются специализированными ферментами. У человека такие ферменты представлены, например, цитохром-оксидазами [92] , глюкуронилтрансферазой [93] , и глутатион S-трансферазой [94]. Эта система ферментов действует в три этапа: на первой стадии ксенобиотики окисляются, затем происходит конъюгирование водорастворимых групп в молекулы, далее модифицированные водорастворимые ксенобиотики могут быть удалены из клеток и метаболизированы перед их экскрецией.
Описанные реакции играют важную роль в разложении микробами загрязняющих веществ и биоремедиации загрязнённых земель и разливов нефти [95]. Многие подобные реакции протекают при участии многоклеточных организмов, однако, ввиду невероятного разнообразия, микроорганизмы справляются с гораздо более широким спектром ксенобиотиков, чем многоклеточные организмы, и способны даже разрушать стойкие органические загрязнители , например хлорорганические соединения [96].
Связанной с этим проблемой для аэробных организмов является оксидативный стресс [97]. В процессе окислительного фосфорилирования и образования дисульфидных связей при укладке белка образуются активные формы кислорода , например пероксид водорода [98]. Эти повреждающие оксиданты удаляются антиоксидантами , например глутатионом и ферментами каталазой и пероксидазами [99] [].
Живые организмы подчиняются началам термодинамики , которые описывают превращения тепла и работы. Второе начало термодинамики гласит, что в любой изолированной системе энтропия не уменьшается. Хотя может показаться, что невероятная сложность живых организмов противоречит этому закону, жизнь возможна, так как все организмы являются открытыми системами, которые обмениваются веществом и энергией с окружающей средой.
Таким образом, живые системы не находятся в термодинамическом равновесии , но, вместо этого, выступают диссипативной системой , которая поддерживает своё состояние сложной организованности, вызывая большее увеличение энтропии окружающей средой []. В метаболизме клеток это достигается путём сочетания спонтанных процессов катаболизма и неспонтанных процессов анаболизма. В термодинамических условиях метаболизм поддерживает порядок за счёт создания беспорядка [].
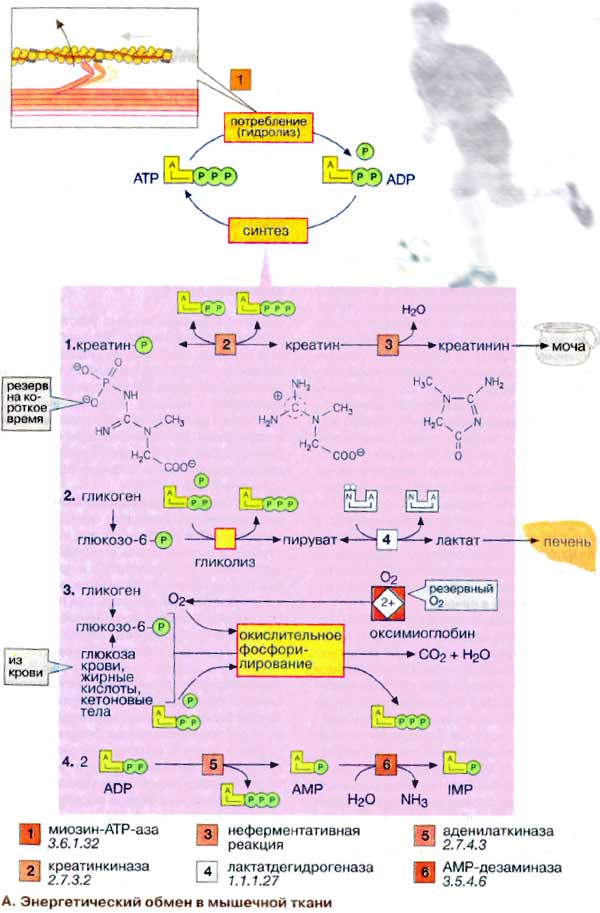
Гомеостазом называют постоянство внутренней среды организма. Так как внешняя среда, окружающая большинство организмов, постоянно меняется, для поддержания постоянных условий внутри клеток реакции обмена веществ должны точно регулироваться [] [].
Регуляция метаболизма позволяет организмам отвечать на сигналы и активно взаимодействовать с окружающей средой []. В случае фермента регуляция заключается в повышении и снижении его активности в ответ на сигналы. С другой стороны, фермент оказывает некоторый контроль над метаболическим путём, который определяется как эффект от изменения активности фермента на данный метаболический путь [].
Выделяют несколько уровней регуляции метаболизма. В метаболическом пути происходит саморегуляция на уровне субстрата или продукта; например, уменьшение количества продукта может компенсированно увеличить поток субстрата реакции по данному пути []. Этот тип регулирования часто включает аллостерическое регулирование активности некоторых ферментов в метаболических путях [].
Внешний контроль включает клетку многоклеточного организма, изменяющую свой метаболизм в ответ на сигналы от других клеток. Эти сигналы, как правило, в виде растворимых мессенджеров, например гормоны и факторы роста , определяются специфическими рецепторами на поверхности клеток []. Затем эти сигналы передаются внутрь клетки системой вторичных мессенджеров , которые зачастую связаны с фосфорилированием белков [].
Хорошо изученный пример внешнего контроля — регуляция метаболизма глюкозы инсулином []. Инсулин вырабатывается в ответ на повышение уровня глюкозы в крови. Гормон связывается с инсулиновым рецептором на поверхности клетки, затем активируется каскад протеинкиназ , которые обеспечивают поглощение молекул глюкозы клетками и преобразовывают их в молекулы жирных кислот и гликогена []. Метаболизм гликогена контролируется активностью фосфорилазы фермента, который расщепляет гликоген и гликогенсинтазы фермента, который образует его.
Эти ферменты взаимосвязаны; фосфорилирование ингибируется гликогенсинтазой, но активируется фосфорилазой. Инсулин вызывает синтез гликогена путём активации белковых фосфатаз и уменьшает фосфорилирование этих ферментов []. Главные пути метаболизма, описанные выше, например гликолиза и цикла Кребса, присутствуют у всех трёх доменах живых существ и обнаруживаются у последнего универсального общего предка [3] [].
Этот универсальный предок был прокариотом и, вероятно, метаногеном с аминокислотным, нуклеотидным, углеводным метаболизмом [] []. Сохранение этих древних метаболических путей в эволюции может быть результатом того, что эти реакции оптимальны для решения конкретных проблем с метаболизмом. Так, конечные продукты гликолиза и цикла Кребса образуются с высокой эффективностью и с минимальным количеством стадий [4] [5].
Первые метаболические пути на основе ферментов могли быть частями пуринового метаболизма нуклеотидов с предыдущим метаболических путей и были частью древнего мира РНК []. Многие модели были предложены для описания механизмов, посредством которых новые метаболические пути эволюционировали.
К ним относятся последовательное добавление новых ферментов на короткий предковый путь, дупликация, а затем дивергенция всех путей, а также набор уже существующих ферментов и их сборка в новый путь реакций []. Относительная важность этих механизмов неясна, однако геномные исследования показали, что ферменты в метаболическом пути, скорее всего, имеют общее происхождение, если предполагать, что многие пути эволюционировали шаг за шагом с новыми функциями, созданными из уже существующих этапов пути [].
Альтернативная модель основана на исследованиях, в которых прослеживается эволюция структуры белков в метаболических связях; предполагают, что ферменты собирались для выполнения схожих функций в различных метаболических путях []. Эти процессы сборки привели к эволюционированию ферментативной мозаики []. Некоторые части обмена веществ, возможно, существовали в качестве «модулей», которые могли быть повторно использованы в различных путях для выполнения схожих функций [].
Эволюция также может приводить к потере метаболических функций. Например, у некоторых паразитов метаболические процессы, которые не важны для выживания, утрачены и готовые аминокислоты, нуклеотиды и углеводы получаются от хозяина []. Подобные упрощения метаболических возможностей наблюдают у эндосимбиотических организмов []. Классически, метаболизм изучается упрощённым подходом, который фокусируется на одном метаболическом пути. Особенно ценно использование меченых атомов на организменном, тканевом и клеточном уровнях, которые определяют пути от предшественников до конечных продуктов путём выявления радиоактивно меченых промежуточных продуктов [].
Ферменты, которые катализируют эти химические реакции, могут затем быть выделены для исследования их кинетики и ответа на ингибиторы. Параллельный подход заключается в выявлении небольших молекул в клетки или ткани; полный набор этих молекул называется метаболом. В целом, эти исследования дают хорошее представление о структуре и функциях простых путей метаболизма, но недостаточны в применении к более сложных системам, например полной метаболизм клетки [].
Идея сложности метаболических сетей в клетках, которые содержат тысячи различных ферментов, отражена на изображении справа, показывающем взаимодействия только между 43 белками и 40 метаболитами, которые регулируются 45 генов []. Тем не менее сейчас можно использовать такие данные о геномах для воссоздания полной сети биохимических реакций и образовывать более целостные математические модели, которые могут объяснить и предсказать их поведение [].
Эти модели особенно сильны, когда используются для интеграции данных о путях и метаболитах, полученных на основе классических методов, с данными по экспрессии генов из протеомных и ДНК-микрочиповых исследований []. С помощью этих методов в настоящее время создаётся модель человеческого метаболизма, которая будет служить ориентиром для будущих исследований лекарств и биохимических исследований [].
Эти модели в настоящее время используются в анализах сети для классификации болезней человека по группам, которые различаются по общим белкам или метаболитам [] []. Яркий пример бактериальных метаболических сетей — устройство галстук-бабочки [] [] [] , структура которой позволяет вводить широкий спектр питательных веществ и производить большое разнообразие продуктов и сложных макромолекул, используя сравнительно немного общих промежуточных веществ.
Основная технологическая основа этой информации — метаболическая инженерия. Здесь организмы, например дрожжи , растения или бактерии, генетически модифицируются, чтобы сделать их более эффективными в биотехнологии и помочь в производстве лекарств, например антибиотиков или промышленных химических веществ, таких как 1,3-пропандиола и шикимовой кислоты [].
Эти генетические модификации обычно направлены на уменьшение количества энергии, используемой для производства продукции, повышения урожайности и снижения производственных отходов []. Термин «метаболизм» впервые введён был в биологию Теодором Шванном в х годах, однако не получил широкого использования. Термин устоялся в физиологии и проник в большинство языков после издания и перевода руководства по физиологии Фостера в х годах [].

История изучения метаболизма охватывает несколько столетий. Исследования начинались с изучения организмов животных, в современной биохимии изучают отдельные метаболические реакции.
Понятие обмена веществ впервые встречается в работах Ибн аль-Нафиса — , который писал, что «тело и его части находятся в постоянном состоянии распада и питания, так что оно неизбежно претерпевает постоянные изменения» []. Первые контролируемые эксперименты по метаболизму у человека были опубликованы Санторио Санторио в году в книге итал. Ars de statica medicina []. Он рассказал, как он сам взвесил себя до и после приёма пищи, сна , работы, секса, натощак, после питья и выделения мочи.
Он обнаружил, что большая часть пищи, которую он принял, была утрачена в результате процесса, названного «незаметным испарением». В ранних исследованиях механизмы метаболических реакций не были обнаружены и считалось, что живой тканью управляет живая сила []. В XIX веке при исследовании ферментации сахара спирта дрожжами Луи Пастер сделал вывод, что брожение катализируется веществами из дрожжевых клеток, которые он назвал ферментами.
Пастер писал, что «алкогольное брожение — действие, связанное с жизнью и организуется дрожжевыми клетками, не связано со смертью или разложением клеток» []. Это открытие, вместе с публикацией Фридриха Вёлера в году о химическом синтезе мочевины [] , доказало, что органические соединения и химические реакции, обнаруженные в клетках, не имеют различий в принципе, как и любые другие разделы химии.
Открытие ферментов в начале XX века Эдуардом Бухнером разделило изучение метаболических реакций от изучения клеток и дало начало развитию биохимии как науки []. Одним из успешных биохимиков начала двадцатого века был Ханс Адольф Кребс , который внёс огромный вклад в изучение метаболизма []. Кребс описал цикл мочевины и позднее, работая вместе с Хансом Корнбергом , цикл лимонной кислоты и глиоксилатный цикл [68] [].
В современных биохимических исследованиях широко используют новые методы, такие как хроматография , рентгеноструктурный анализ , ЯМР-спектроскопия , электронная микроскопия и метод классической молекулярной динамики. Эти методы позволяют открывать и подробно изучать множество молекул и метаболических путей в клетках. Материал из Википедии — свободной энциклопедии. Запрос «Метаболизм» перенаправляется сюда; об архитектурной концепции см. Метаболизм архитектура. Основная статья: Белки.
Основные статьи: Липиды и Липидный обмен. Основные статьи: Углеводы и Углеводный обмен. Основная статья: Нуклеотиды. Подробное рассмотрение темы: Коферменты. Основная статья: Бионеорганическая химия. Основная статья: Катаболизм.
Подробное рассмотрение темы: Пищеварение и Желудочно-кишечный тракт. Подробное рассмотрение темы: Клеточное дыхание , Брожение , Липолиз и Бета-окисление. Подробное рассмотрение темы: Окислительное фосфорилирование , Хемиосмос и Митохондрия. Подробное рассмотрение темы: Анаболизм. Подробное рассмотрение темы: Фотосинтез и Хемосинтез. Подробное рассмотрение темы: Глюконеогенез и Гликозилирование.
Подробное рассмотрение темы: Стероиды и Биосинтез жирных кислот. Подробное рассмотрение темы: Биосинтез белка. Подробное рассмотрение темы: Пурин и пиримидин. Подробное рассмотрение темы: Антиоксиданты. Подробное рассмотрение темы: Гормоны и Передача сигнала в клетке. Подробное рассмотрение темы: Филогенетика. Подробное рассмотрение темы: Протеомика и Метабономика.
Подробное рассмотрение темы: История биохимии и История молекулярной биологии. Physiology and genetics of sulfur-oxidizing bacteria англ. The universal nature of biochemistry англ. Universality in intermediary metabolism англ.
Архивировано 2 июня года. Evolutionary optimization of metabolic pathways. The puzzle of the Krebs citric acid cycle: assembling the pieces of chemically feasible reactions, and opportunism in the design of metabolic pathways during evolution англ. Dynamic filaments of the bacterial cytoskeleton англ. Lehninger Principles of Biochemistry неопр.
Freeman and company, Kelleher, B. Bryan, R. Mallet, A. Holleran, A. Архивировано 22 сентября года. Hothersall, Aamir Ahmed. A comprehensive classification system for lipids англ. Архивировано 24 августа года. Дата обращения: 8 марта Архивировано 22 августа года. Mitochondrial 3-hydroxymethylglutaryl-CoA synthase: a control enzyme in ketogenesis англ.
Glycomics: an integrated systems approach to structure-function relationships of glycans англ. Basics of the virology of HIV-1 and its replication англ.
Mechanisms of enzyme-catalyzed group transfer reactions англ. Compartmentation and communication in living systems. Ligand conduction: a general catalytic principle in chemical, osmotic and chemiosmotic reaction systems англ. Catalytic and mechanical cycles in F-ATP synthases.

Fourth in the Cycles Review Series англ. Stanford School of Medicine Nutrition Courses англ. The power to reduce: pyridine nucleotides—small molecules with a multitude of functions англ. Chemical and elemental analysis of humans in vivo using improved body composition models англ.
Yeast as a model organism to study transport and homeostasis of alkali metal cations англ. Архивировано 25 октября года. Modulation of ion channels in neurons and other cells англ.
Excitation-contraction coupling from the s into the new millennium англ. Macro- and micromineral composition of pigs from birth to kilograms of body weight англ. Архивировано 30 апреля года. Elemental fingerprint analysis of barley Hordeum vulgare using inductively coupled plasma mass spectrometry, isotope-ratio mass spectrometry, and multivariate statistics англ.
Transition metal speciation in the cell: insights from the chemistry of metal ion receptors англ. Mammalian zinc transport, trafficking, and signals англ. Архивировано 5 ноября года. Iron uptake and metabolism in the new millennium англ. Нетрусов, И. Котова — М.
Гусев, Л. Минеева — 4-е изд. Биология прокариотов: Учебник. Том Lwoff, C. Ryan et al. Nomenclature of nutritional types of microorganisms. Архивировано 7 ноября года. Life: past, present and future англ. Архивировано 17 марта года. The complex architecture of oxygenic photosynthesis англ. Bacterial extracellular zinc-containing metalloproteases рум. Bacterial lipases: an overview of production, purification and biochemical properties англ. The digestive system: linking theory and practice неопр.
How amino acids get into cells: mechanisms, models, menus, and mediators англ. Structure and function of facilitative sugar transporters англ.
Structure and function of mammalian facilitative sugar transporters англ. The cellular fate of glucose and its relevance in type 2 diabetes англ. Архивировано 4 декабря года. Amino acid metabolism англ. Glutamate, at the interface between amino acid and carbohydrate metabolism англ. Архивировано 28 февраля года. Glutamine: the emperor or his clothes? Архивировано 30 ноября года.
Energy transduction: proton transfer through the respiratory complexes англ. Structures and proton-pumping strategies of mitochondrial respiratory enzymes англ. Molecular biology of hydrogen utilization in aerobic chemolithotrophs англ.
Microorganisms pumping iron: anaerobic microbial iron oxidation and reduction англ. The anaerobic oxidation of ammonium неопр.
Enzymology and bioenergetics of respiratory nitrite ammonification англ. Microbial co-operation in the rhizosphere англ. Архивировано 2 декабря года. Diel variations in carbon metabolism by green nonsulfur-like bacteria in alkaline siliceous hot spring microbial mats from Yellowstone National Park англ. Interactive control of Rhodobacter capsulatus redox-balancing systems during phototrophic metabolism англ. Photosynthetic reaction centers англ. Cyclic electron flow around photosystem I is essential for photosynthesis англ.
Ribulose-1,5-bisphosphate carboxylase-oxygenase англ. Crassulacean acid metabolism: plastic, fantastic англ. Архивировано 25 января года. Evidence for autotrophic CO 2 fixation via the reductive tricarboxylic acid cycle by members of the epsilon subdivision of proteobacteria англ.
Enzymes of a novel autotrophic CO 2 fixation pathway in the phototrophic bacterium Chloroflexus aurantiacus, the 3-hydroxypropionate cycle англ. Архивировано 5 августа года. Something from almost nothing: carbon dioxide fixation in chemoautotrophs англ. Design of glycolysis англ. Fructose-2,6-bisphosphate in control of hepatic gluconeogenesis. From metabolites to molecular genetics англ. Revisiting the glyoxylate cycle: alternate pathways for microbial acetate assimilation англ.
Proteolytic and lipolytic responses to starvation неопр. Synthesis of cell constituents from C2-units by a modified tricarboxylic acid cycle англ. Evans, Lisa C. Metabolic pathways and abnormalities англ. Архивировано 11 августа года.
Freeze, Gerald W. Hart, Ronald L. Cummings, Jeffrey D. Esko, Pamela Stanley, Gerald W. Hart, Markus Aebi, Alan G. Darvill, Taroh Kinoshita, Nicolle H. Packer, James H.